You are browsing environment: HUMAN GUT
CAZyme Information: MGYG000004797_00483
You are here: Home > Sequence: MGYG000004797_00483
Basic Information |
Genomic context |
Full Sequence |
Enzyme annotations |
CAZy signature domains |
CDD domains |
CAZyme hits |
PDB hits |
Swiss-Prot hits |
SignalP and Lipop annotations |
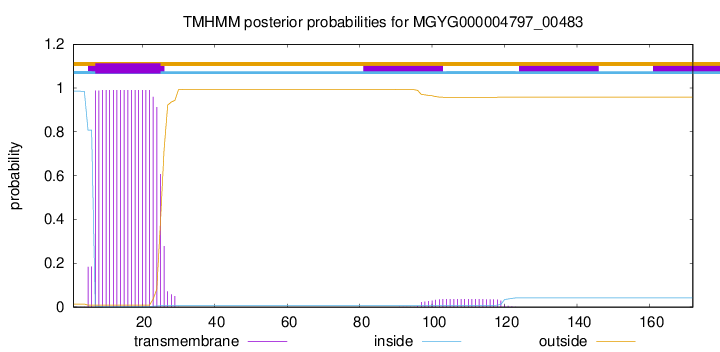
TMHMM annotations
Basic Information help
| Species | Phocaeicola sartorii | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Lineage | Bacteria; Bacteroidota; Bacteroidia; Bacteroidales; Bacteroidaceae; Phocaeicola; Phocaeicola sartorii | |||||||||||
| CAZyme ID | MGYG000004797_00483 | |||||||||||
| CAZy Family | GH24 | |||||||||||
| CAZyme Description | hypothetical protein | |||||||||||
| CAZyme Property |
|
|||||||||||
| Genome Property |
|
|||||||||||
| Gene Location | Start: 95061; End: 95579 Strand: - | |||||||||||
CAZyme Signature Domains help
| Family | Start | End | Evalue | family coverage |
|---|---|---|---|---|
| GH24 | 49 | 163 | 1.6e-16 | 0.8394160583941606 |
CDD Domains download full data without filtering help
| Cdd ID | Domain | E-Value | qStart | qEnd | sStart | sEnd | Domain Description |
|---|---|---|---|---|---|---|---|
| cd00737 | lyz_endolysin_autolysin | 6.46e-20 | 37 | 166 | 1 | 134 | endolysin and autolysin. The dsDNA phages of eubacteria use endolysins or muralytic enzymes in conjunction with hollin, a small membrane protein, to degrade the peptidoglycan found in bacterial cell walls. Similarly, bacteria produce autolysins to facilitate the biosynthesis of its cell wall heteropolymer peptidoglycan and cell division. Endolysins and autolysins are found in viruses and bacteria, respectively. Both endolysin and autolysin enzymes cleave the glycosidic beta 1,4-bonds between the N-acetylmuramic acid and the N-acetylglucosamine of the peptidoglycan. |
| COG3772 | RrrD | 1.14e-12 | 34 | 170 | 8 | 150 | Phage-related lysozyme (muramidase), GH24 family [Cell wall/membrane/envelope biogenesis]. |
| cd16900 | endolysin_R21-like | 5.42e-08 | 54 | 166 | 30 | 140 | endolysin R21-like proteins. Unlike T4 E phage lysozyme, the endolysin R21 from Enterobacteria phage P21 has an N-terminal SAR (signal-arrest-release) domain that anchors the endolysin to the membrane in an inactive form, which act to prevent premature lysis of the infected bacterium. The dsDNA phages of eubacteria use endolysins or muralytic enzymes in conjunction with hollin, a small membrane protein, to degrade the peptidoglycan found in bacterial cell walls. Similarly, bacteria produce autolysins to facilitate the biosynthesis of its cell wall heteropolymer peptidoglycan and cell division. Endolysins and autolysins are found in viruses and bacteria, respectively. Both endolysin and autolysin enzymes cleave the glycosidic beta 1,4-bonds between the N-acetylmuramic acid and the N-acetylglucosamine of the peptidoglycan. |
| cd00735 | T4-like_lys | 0.002 | 51 | 141 | 21 | 117 | bacteriophage T4-like lysozymes. Bacteriophage T4-like lysozymes hydrolyze the beta-1,4-glycosidic bond between N-acetylmuramic acid (MurNAc) and N-acetylglucosamine (GlcNAc) in peptidoglycan heteropolymers of prokaryotic cell walls. Members include a variety of bacteriophages (T4, RB49, RB69, Aeh1), as well as Dictyostelium. |
CAZyme Hits help
| Hit ID | E-Value | Query Start | Query End | Hit Start | Hit End |
|---|---|---|---|---|---|
| QCD36945.1 | 4.50e-65 | 2 | 167 | 1 | 160 |
| QCP71761.1 | 4.52e-55 | 2 | 167 | 1 | 159 |
| QCD40659.1 | 4.52e-55 | 2 | 167 | 1 | 159 |
| QCD38079.1 | 4.52e-55 | 2 | 167 | 1 | 159 |
| QCP71601.1 | 1.02e-51 | 2 | 167 | 1 | 159 |
Swiss-Prot Hits help
SignalP and Lipop Annotations help
This protein is predicted as SP
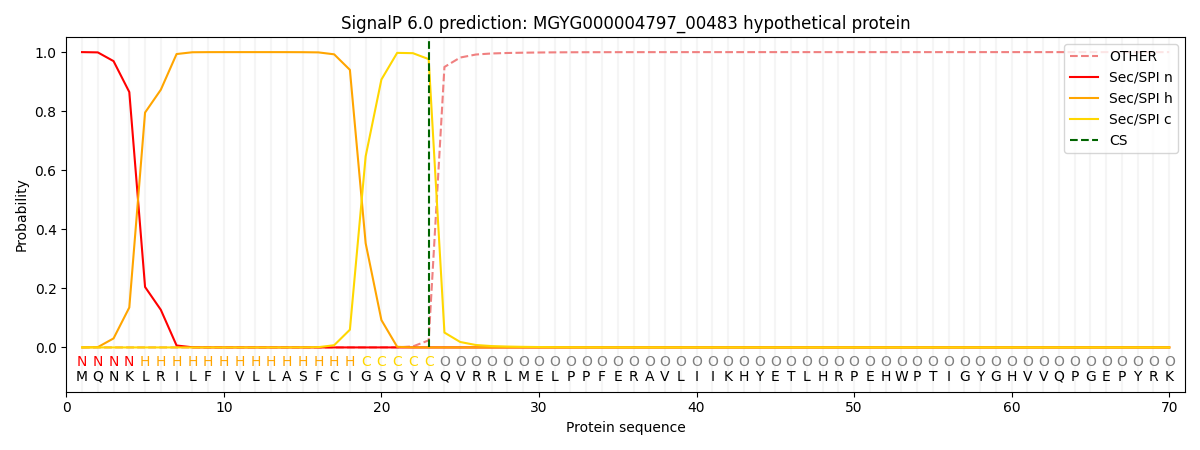
| Other | SP_Sec_SPI | LIPO_Sec_SPII | TAT_Tat_SPI | TATLIP_Sec_SPII | PILIN_Sec_SPIII |
|---|---|---|---|---|---|
| 0.000457 | 0.998663 | 0.000393 | 0.000162 | 0.000149 | 0.000141 |