You are browsing environment: HUMAN GUT
CAZyme Information: MGYG000002717_02909
You are here: Home > Sequence: MGYG000002717_02909
Basic Information |
Genomic context |
Full Sequence |
Enzyme annotations |
CAZy signature domains |
CDD domains |
CAZyme hits |
PDB hits |
Swiss-Prot hits |
SignalP and Lipop annotations |
TMHMM annotations
Basic Information help
| Species | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Lineage | Bacteria; Bacteroidota; Bacteroidia; Bacteroidales; Bacteroidaceae; Bacteroides; | |||||||||||
| CAZyme ID | MGYG000002717_02909 | |||||||||||
| CAZy Family | GH43 | |||||||||||
| CAZyme Description | Extracellular endo-alpha-(1->5)-L-arabinanase | |||||||||||
| CAZyme Property |
|
|||||||||||
| Genome Property |
|
|||||||||||
| Gene Location | Start: 3505; End: 4992 Strand: - | |||||||||||
CAZyme Signature Domains help
| Family | Start | End | Evalue | family coverage |
|---|---|---|---|---|
| GH43 | 53 | 365 | 9.2e-98 | 0.9967741935483871 |
CDD Domains download full data without filtering help
| Cdd ID | Domain | E-Value | qStart | qEnd | sStart | sEnd | Domain Description |
|---|---|---|---|---|---|---|---|
| cd08998 | GH43_Arb43a-like | 1.10e-106 | 54 | 360 | 1 | 278 | Glycosyl hydrolase family 43 protein such as Bacillus subtilis subsp. subtilis str. 168 endo-alpha-1,5-L-arabinanase Arb43A. This glycosyl hydrolase family 43 (GH43) subgroup belongs to the glycosyl hydrolase clan F (according to carbohydrate-active enzymes database (CAZY)) which includes family 43 (GH43) and 62 (GH62) families. GH43 are inverting enzymes (i.e. they invert the stereochemistry of the anomeric carbon atom of the substrate) that have an aspartate as the catalytic general base, a glutamate as the catalytic general acid and another aspartate that is responsible for pKa modulation and orienting the catalytic acid. The GH43 ABN enzymes hydrolyze alpha-1,5-L-arabinofuranoside linkages while the ABF enzymes cleave arabinose side chains so that the combined actions of these two enzymes reduce arabinan to L-arabinose and/or arabinooligosaccharides. Many of these enzymes such as the Bacillus subtilis arabinanase Abn2, that hydrolyzes sugar beet arabinan (branched), linear alpha-1,5-L-arabinan and pectin, are different from other arabinases; they are organized into two different domains with a divalent metal cluster close to the catalytic residues to guarantee the correct protonation state of the catalytic residues and consequently the enzyme activity. These arabinan-degrading enzymes are important in the food industry for efficient production of L-arabinose from agricultural waste; L-arabinose is often used as a bioactive sweetener. A common structural feature of GH43 enzymes is a 5-bladed beta-propeller domain that contains the catalytic acid and catalytic base. A long V-shaped groove, partially enclosed at one end, forms a single extended substrate-binding surface across the face of the propeller. |
| cd18830 | GH43_CjArb43A-like | 3.56e-52 | 54 | 339 | 1 | 271 | Glycosyl hydrolase family 43 protein such as Cellvibrio japonicus Ueda107 endo-alpha-1,5-L-arabinanase / exo-alpha-1,5-L-arabinanase 43A (ArbA;CJA_0805) (Arb43A). This glycosyl hydrolase family 43 (GH43) subgroup includes mostly enzymes annotated with alpha-L-arabinofuranosidase (ABF; EC 3.2.1.55) and endo-alpha-L-arabinanase (ABN; EC 3.2.1.99) activities, and includes the bifunctional Cellvibrio japonicus Ueda107 endo-alpha-1,5-L-arabinanase / exo-alpha-1,5-L-arabinanase 43A (ArbA;CJA_0805) (Arb43A). It belongs to the glycosyl hydrolase clan F (according to carbohydrate-active enzymes database (CAZY)) which includes family 43 (GH43) and 62 (GH62) families. GH43 are inverting enzymes (i.e. they invert the stereochemistry of the anomeric carbon atom of the substrate) that have an aspartate as the catalytic general base, a glutamate as the catalytic general acid and another aspartate that is responsible for pKa modulation and orienting the catalytic acid. The GH43 ABN enzymes hydrolyze alpha-1,5-L-arabinofuranoside linkages while the ABF enzymes cleave arabinose side chains so that the combined actions of these two enzymes reduce arabinan to L-arabinose and/or arabinooligosaccharides. Many of these enzymes such as the Bacillus subtilis arabinanase Abn2, that hydrolyzes sugar beet arabinan (branched), linear alpha-1,5-L-arabinan and pectin, are different from other arabinases; they are organized into two different domains with a divalent metal cluster close to the catalytic residues to guarantee the correct protonation state of the catalytic residues and consequently the enzyme activity. These arabinan-degrading enzymes are important in the food industry for efficient production of L-arabinose from agricultural waste; L-arabinose is often used as a bioactive sweetener. A common structural feature of GH43 enzymes is a 5-bladed beta-propeller domain that contains the catalytic acid and catalytic base. A long V-shaped groove, partially enclosed at one end, forms a single extended substrate-binding surface across the face of the propeller. |
| cd18616 | GH43_ABN-like | 1.99e-51 | 56 | 337 | 10 | 263 | Glycosyl hydrolase family 43 such as arabinan endo-1 5-alpha-L-arabinosidase. This glycosyl hydrolase family 43 (GH43) subgroup includes mostly enzymes with endo-alpha-L-arabinanase (ABN; EC 3.2.1.99) activity. These are inverting enzymes (i.e. they invert the stereochemistry of the anomeric carbon atom of the substrate) that have an aspartate as the catalytic general base, a glutamate as the catalytic general acid and another aspartate that is responsible for pKa modulation and orienting the catalytic acid. The GH43 ABN enzymes hydrolyze alpha-1,5-L-arabinofuranoside linkages. These arabinan-degrading enzymes are important in the food industry for efficient production of L-arabinose from agricultural waste; L-arabinose is often used as a bioactive sweetener. A common structural feature of GH43 enzymes is a 5-bladed beta-propeller domain that contains the catalytic acid and catalytic base. A long V-shaped groove, partially enclosed at one end, forms a single extended substrate-binding surface across the face of the propeller. |
| cd18832 | GH43_GsAbnA-like | 4.63e-50 | 54 | 360 | 1 | 332 | Glycosyl hydrolase family 43 protein such as Geobacillus stearothermophilus endo-alpha-1,5-L-arabinanase AbnA. This glycosyl hydrolase family 43 (GH43) subgroup includes mostly enzymes with alpha-L-arabinofuranosidase (ABF; EC 3.2.1.55) and endo-alpha-L-arabinanase (ABN; EC 3.2.1.99) activities. It includes Geobacillus stearothermophilus T-6 NCIMB 40222 AbnA, Bacillus subtilis subsp. subtilis str. 168 (Abn2;YxiA;J3A;BSU39330) (Arb43B), and Thermotoga petrophila RKU-1 (AbnA;TpABN;Tpet_0637). These are inverting enzymes (i.e. they invert the stereochemistry of the anomeric carbon atom of the substrate) that have an aspartate as the catalytic general base, a glutamate as the catalytic general acid and another aspartate that is responsible for pKa modulation and orienting the catalytic acid. The GH43 ABN enzymes hydrolyze alpha-1,5-L-arabinofuranoside linkages while the ABF enzymes cleave arabinose side chains so that the combined actions of these two enzymes reduce arabinan to L-arabinose and/or arabinooligosaccharides. Many of these enzymes are different from other arabinases; they are organized into two different domains with a divalent metal cluster close to the catalytic residues to guarantee the correct protonation state of the catalytic residues and consequently the enzyme activity. These arabinan-degrading enzymes are important in the food industry for efficient production of L-arabinose from agricultural waste; L-arabinose is often used as a bioactive sweetener. A common structural feature of GH43 enzymes is a 5-bladed beta-propeller domain that contains the catalytic acid and catalytic base. A long V-shaped groove, partially enclosed at one end, forms a single extended substrate-binding surface across the face of the propeller. |
| cd08988 | GH43_ABN | 3.81e-46 | 55 | 358 | 1 | 276 | Glycosyl hydrolase family 43. This glycosyl hydrolase family 43 (GH43) subgroup includes mostly enzymes with alpha-L-arabinofuranosidase (ABF; EC 3.2.1.55) and endo-alpha-L-arabinanase (ABN; EC 3.2.1.99) activities. These are inverting enzymes (i.e. they invert the stereochemistry of the anomeric carbon atom of the substrate) that have an aspartate as the catalytic general base, a glutamate as the catalytic general acid and another aspartate that is responsible for pKa modulation and orienting the catalytic acid. The GH43 ABN enzymes hydrolyze alpha-1,5-L-arabinofuranoside linkages while the ABF enzymes cleave arabinose side chains so that the combined actions of these two enzymes reduce arabinan to L-arabinose and/or arabinooligosaccharides. These arabinan-degrading enzymes are important in the food industry for efficient production of L-arabinose from agricultural waste; L-arabinose is often used as a bioactive sweetener. A common structural feature of GH43 enzymes is a 5-bladed beta-propeller domain that contains the catalytic acid and catalytic base. A long V-shaped groove, partially enclosed at one end, forms a single extended substrate-binding surface across the face of the propeller. |
CAZyme Hits help
| Hit ID | E-Value | Query Start | Query End | Hit Start | Hit End |
|---|---|---|---|---|---|
| QDO70910.1 | 0.0 | 1 | 495 | 1 | 495 |
| QRQ48074.1 | 0.0 | 1 | 495 | 1 | 495 |
| QUT46362.1 | 0.0 | 1 | 495 | 1 | 495 |
| QUT90798.1 | 1.02e-317 | 1 | 495 | 1 | 495 |
| ALJ58184.1 | 4.09e-317 | 1 | 495 | 1 | 495 |
PDB Hits download full data without filtering help
| Hit ID | E-Value | Query Start | Query End | Hit Start | Hit End | Description |
|---|---|---|---|---|---|---|
| 4KC7_A | 6.65e-53 | 38 | 495 | 19 | 473 | CrystalStructure of Endo-1,5-alpha-L-arabinanase from Thermotoga petrophila RKU-1 [Thermotoga petrophila RKU-1],4KC7_B Crystal Structure of Endo-1,5-alpha-L-arabinanase from Thermotoga petrophila RKU-1 [Thermotoga petrophila RKU-1],4KC7_C Crystal Structure of Endo-1,5-alpha-L-arabinanase from Thermotoga petrophila RKU-1 [Thermotoga petrophila RKU-1],4KC8_A Crystal Structure of Endo-1,5-alpha-L-arabinanase from Thermotoga petrophila RKU-1 in complex with TRIS [Thermotoga petrophila RKU-1],4KC8_B Crystal Structure of Endo-1,5-alpha-L-arabinanase from Thermotoga petrophila RKU-1 in complex with TRIS [Thermotoga petrophila RKU-1],4KC8_C Crystal Structure of Endo-1,5-alpha-L-arabinanase from Thermotoga petrophila RKU-1 in complex with TRIS [Thermotoga petrophila RKU-1] |
| 2X8F_A | 7.69e-43 | 53 | 493 | 35 | 469 | Nativestructure of Endo-1,5-alpha-L-arabinanases from Bacillus subtilis [Bacillus subtilis],2X8F_B Native structure of Endo-1,5-alpha-L-arabinanases from Bacillus subtilis [Bacillus subtilis] |
| 2X8T_A | 2.07e-42 | 53 | 493 | 35 | 469 | CrystalStructure of the Abn2 H318A mutant [Bacillus subtilis],2X8T_B Crystal Structure of the Abn2 H318A mutant [Bacillus subtilis] |
| 2X8S_A | 1.02e-41 | 53 | 493 | 35 | 469 | CrystalStructure of the Abn2 D171A mutant in complex with arabinotriose [Bacillus subtilis],2X8S_B Crystal Structure of the Abn2 D171A mutant in complex with arabinotriose [Bacillus subtilis] |
| 4COT_A | 1.38e-41 | 53 | 493 | 35 | 469 | Theimportance of the Abn2 calcium cluster in the endo-1,5- arabinanase activity from Bacillus subtilis [Bacillus subtilis subsp. subtilis str. 168] |
Swiss-Prot Hits download full data without filtering help
| Hit ID | E-Value | Query Start | Query End | Hit Start | Hit End | Description |
|---|---|---|---|---|---|---|
| A5IKD4 | 4.76e-52 | 54 | 495 | 30 | 470 | Extracellular endo-alpha-(1->5)-L-arabinanase OS=Thermotoga petrophila (strain ATCC BAA-488 / DSM 13995 / JCM 10881 / RKU-1) OX=390874 GN=Tpet_0637 PE=1 SV=1 |
| P42293 | 4.14e-42 | 53 | 493 | 35 | 469 | Extracellular endo-alpha-(1->5)-L-arabinanase 2 OS=Bacillus subtilis (strain 168) OX=224308 GN=abn2 PE=1 SV=2 |
| B3EYM8 | 1.14e-39 | 54 | 368 | 25 | 314 | Intracellular endo-alpha-(1->5)-L-arabinanase OS=Geobacillus stearothermophilus OX=1422 GN=abnB PE=1 SV=1 |
| Q93HT9 | 2.85e-37 | 54 | 365 | 25 | 311 | Intracellular endo-alpha-(1->5)-L-arabinanase OS=Geobacillus thermodenitrificans OX=33940 GN=abn-ts PE=1 SV=1 |
| P95470 | 2.36e-31 | 53 | 367 | 35 | 333 | Extracellular exo-alpha-(1->5)-L-arabinofuranosidase ArbA OS=Cellvibrio japonicus (strain Ueda107) OX=498211 GN=arbA PE=1 SV=1 |
SignalP and Lipop Annotations help
This protein is predicted as LIPO
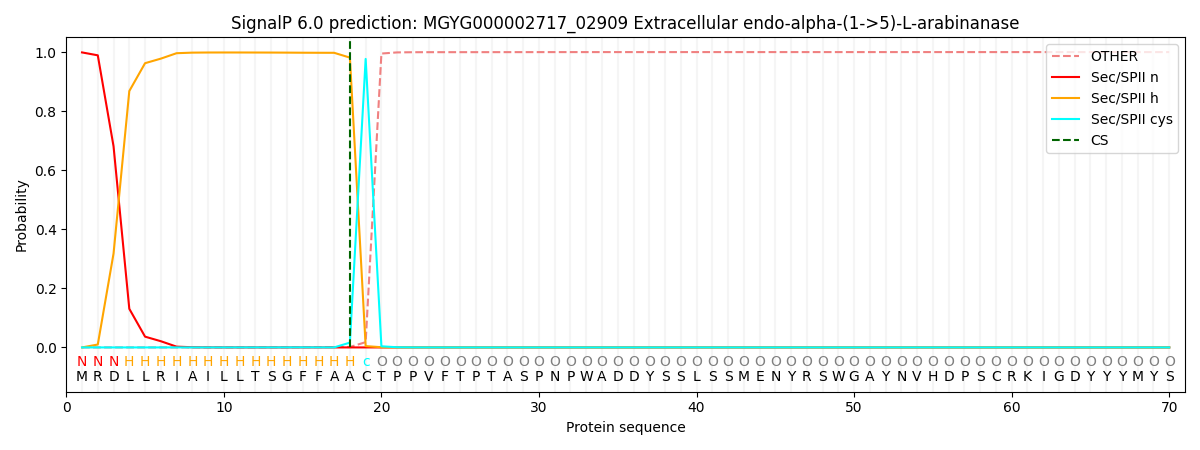
| Other | SP_Sec_SPI | LIPO_Sec_SPII | TAT_Tat_SPI | TATLIP_Sec_SPII | PILIN_Sec_SPIII |
|---|---|---|---|---|---|
| 0.000005 | 0.001090 | 0.998939 | 0.000001 | 0.000001 | 0.000001 |