You are browsing environment: HUMAN GUT
CAZyme Information: MGYG000004739_01797
You are here: Home > Sequence: MGYG000004739_01797
Basic Information |
Genomic context |
Full Sequence |
Enzyme annotations |
CAZy signature domains |
CDD domains |
CAZyme hits |
PDB hits |
Swiss-Prot hits |
SignalP and Lipop annotations |
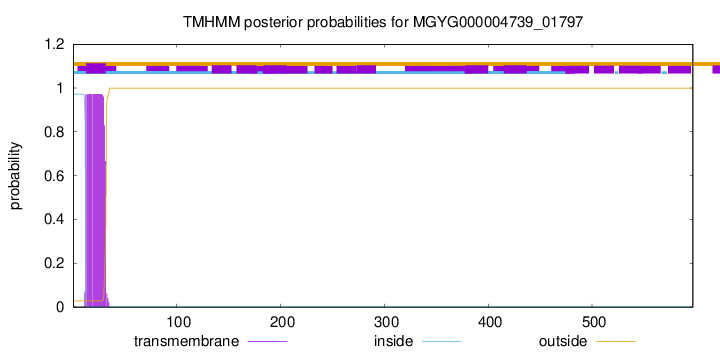
TMHMM annotations
Basic Information help
| Species | Actinomyces sp000220835 | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Lineage | Bacteria; Actinobacteriota; Actinomycetia; Actinomycetales; Actinomycetaceae; Actinomyces; Actinomyces sp000220835 | |||||||||||
| CAZyme ID | MGYG000004739_01797 | |||||||||||
| CAZy Family | GH68 | |||||||||||
| CAZyme Description | Levansucrase | |||||||||||
| CAZyme Property |
|
|||||||||||
| Genome Property |
|
|||||||||||
| Gene Location | Start: 6786; End: 8579 Strand: + | |||||||||||
CAZyme Signature Domains help
| Family | Start | End | Evalue | family coverage |
|---|---|---|---|---|
| GH68 | 91 | 571 | 1.2e-146 | 0.9832134292565947 |
CDD Domains download full data without filtering help
| Cdd ID | Domain | E-Value | qStart | qEnd | sStart | sEnd | Domain Description |
|---|---|---|---|---|---|---|---|
| pfam02435 | Glyco_hydro_68 | 3.27e-139 | 91 | 566 | 1 | 410 | Levansucrase/Invertase. This Pfam family consists of the glycosyl hydrolase 68 family, including several bacterial levansucrase enzymes, and invertase from zymomonas. |
| cd08997 | GH68 | 1.66e-117 | 141 | 565 | 1 | 354 | Glycosyl hydrolase family 68, includes levansucrase, beta-fructofuranosidase and inulosucrase. Glycosyl hydrolase family 68 (GH68) consists of frucosyltransferases (FTFs) that include levansucrase (EC 2.4.1.10), beta-fructofuranosidase (EC 3.2.1.26) and inulosucrase (EC 2.4.1.9), all of which use sucrose as their preferential donor substrate. Levansucrase, also known as beta-D-fructofuranosyl transferase, catalyzes the transfer of the sucrose fructosyl moiety to a growing levan chain. Similarly, inulosucrase catalyzes long inulin-type of fructans, and beta-fructofuranosidases create fructooligosaccharides (FOS). However, in the absence of high fructan/sucrose ratio, some GH68 enzymes can also use fructan as donor substrate. GH68 retaining enzymes (i.e. they retain the configuration at anomeric carbon atom of the substrate) catalyze hydrolysis in two steps involving a covalent glycosyl enzyme intermediate: an aspartate located close to the N-terminus acts as the catalytic nucleophile and a glutamate acts as the general acid/base; a conserved aspartate residue in the Arg-Asp-Pro (RDP) motif stabilizes the transition state. A common structural feature of all these enzymes is a 5-bladed beta-propeller domain, similar to GH43, that contains the catalytic acid and catalytic base. A long V-shaped groove, partially enclosed at one end, forms a single extended substrate-binding surface across the face of the propeller. Biotechnological applications of these enzymes include use of inulin in inexpensive production of rich fructose syrups as well as use of FOS as health-promoting pre-biotics. |
| cd18609 | GH32-like | 7.44e-16 | 137 | 484 | 5 | 258 | Glycosyl hydrolase family 32 family protein. The GH32 family contains glycosyl hydrolase family GH32 proteins that cleave sucrose into fructose and glucose via beta-fructofuranosidase activity, producing invert sugar that is a mixture of dextrorotatory D-glucose and levorotatory D-fructose, thus named invertase (EC 3.2.1.26). This family also contains other fructofuranosidases such as inulinase (EC 3.2.1.7), exo-inulinase (EC 3.2.1.80), levanase (EC 3.2.1.65), and transfructosidases such sucrose:sucrose 1-fructosyltransferase (EC 2.4.1.99), fructan:fructan 1-fructosyltransferase (EC 2.4.1.100), sucrose:fructan 6-fructosyltransferase (EC 2.4.1.10), fructan:fructan 6G-fructosyltransferase (EC 2.4.1.243) and levan fructosyltransferases (EC 2.4.1.-). These retaining enzymes (i.e. they retain the configuration at anomeric carbon atom of the substrate) catalyze hydrolysis in two steps involving a covalent glycosyl enzyme intermediate: an aspartate located close to the N-terminus acts as the catalytic nucleophile and a glutamate acts as the general acid/base; a conserved aspartate residue in the Arg-Asp-Pro (RDP) motif stabilizes the transition state. These enzymes are predicted to display a 5-fold beta-propeller fold as found for GH43 and CH68. The breakdown of sucrose is widely used as a carbon or energy source by bacteria, fungi, and plants. Invertase is used commercially in the confectionery industry, since fructose has a sweeter taste than sucrose and a lower tendency to crystallize. A common structural feature of all these enzymes is a 5-bladed beta-propeller domain, similar to GH43, that contains the catalytic acid and catalytic base. A long V-shaped groove, partially enclosed at one end, forms a single extended substrate-binding surface across the face of the propeller. |
| cd08979 | GH_J | 1.06e-07 | 227 | 467 | 54 | 223 | Glycosyl hydrolase families 32 and 68, which form the clan GH-J. This glycosyl hydrolase family clan J (according to carbohydrate-active enzymes database (CAZY)) includes family 32 (GH32) and 68 (GH68). GH32 enzymes include invertase (EC 3.2.1.26) and other other fructofuranosidases such as inulinase (EC 3.2.1.7), exo-inulinase (EC 3.2.1.80), levanase (EC 3.2.1.65), and transfructosidases such sucrose:sucrose 1-fructosyltransferase (EC 2.4.1.99), fructan:fructan 1-fructosyltransferase (EC 2.4.1.100), sucrose:fructan 6-fructosyltransferase (EC 2.4.1.10), fructan:fructan 6G-fructosyltransferase (EC 2.4.1.243) and levan fructosyltransferases (EC 2.4.1.-). The GH68 family consists of frucosyltransferases (FTFs) that include levansucrase (EC 2.4.1.10, also known as beta-D-fructofuranosyl transferase), beta-fructofuranosidase (EC 3.2.1.26) and inulosucrase (EC 2.4.1.9). GH32 and GH68 family enzymes are retaining enzymes (i.e. they retain the configuration at anomeric carbon atom of the substrate) and catalyze hydrolysis in two steps involving a covalent glycosyl enzyme intermediate: an aspartate located close to the N-terminus acts as the catalytic nucleophile and a glutamate acts as the general acid/base; a conserved aspartate residue in the Arg-Asp-Pro (RDP) motif stabilizes the transition state. A common structural feature of all these enzymes is a 5-bladed beta-propeller domain, similar to GH43, that contains the catalytic acid and catalytic base. A long V-shaped groove, partially enclosed at one end, forms a single extended substrate-binding surface across the face of the propeller. |
| cd08996 | GH32_FFase | 0.006 | 236 | 344 | 64 | 139 | Glycosyl hydrolase family 32, beta-fructosidases. Glycosyl hydrolase family GH32 cleaves sucrose into fructose and glucose via beta-fructofuranosidase activity, producing invert sugar that is a mixture of dextrorotatory D-glucose and levorotatory D-fructose, thus named invertase (EC 3.2.1.26). This family also contains other fructofuranosidases such as inulinase (EC 3.2.1.7), exo-inulinase (EC 3.2.1.80), levanase (EC 3.2.1.65), and transfructosidases such sucrose:sucrose 1-fructosyltransferase (EC 2.4.1.99), fructan:fructan 1-fructosyltransferase (EC 2.4.1.100), sucrose:fructan 6-fructosyltransferase (EC 2.4.1.10), fructan:fructan 6G-fructosyltransferase (EC 2.4.1.243) and levan fructosyltransferases (EC 2.4.1.-). These retaining enzymes (i.e. they retain the configuration at anomeric carbon atom of the substrate) catalyze hydrolysis in two steps involving a covalent glycosyl enzyme intermediate: an aspartate located close to the N-terminus acts as the catalytic nucleophile and a glutamate acts as the general acid/base; a conserved aspartate residue in the Arg-Asp-Pro (RDP) motif stabilizes the transition state. These enzymes are predicted to display a 5-fold beta-propeller fold as found for GH43 and CH68. The breakdown of sucrose is widely used as a carbon or energy source by bacteria, fungi, and plants. Invertase is used commercially in the confectionery industry, since fructose has a sweeter taste than sucrose and a lower tendency to crystallize. A common structural feature of all these enzymes is a 5-bladed beta-propeller domain, similar to GH43, that contains the catalytic acid and catalytic base. A long V-shaped groove, partially enclosed at one end, forms a single extended substrate-binding surface across the face of the propeller. |
CAZyme Hits help
| Hit ID | E-Value | Query Start | Query End | Hit Start | Hit End |
|---|---|---|---|---|---|
| VEG27298.1 | 0.0 | 25 | 597 | 9 | 566 |
| QQQ59919.1 | 0.0 | 1 | 597 | 3 | 636 |
| QQC38808.1 | 0.0 | 62 | 597 | 95 | 636 |
| QLF52964.1 | 0.0 | 62 | 597 | 95 | 636 |
| AMD98227.1 | 0.0 | 62 | 597 | 95 | 636 |
PDB Hits download full data without filtering help
| Hit ID | E-Value | Query Start | Query End | Hit Start | Hit End | Description |
|---|---|---|---|---|---|---|
| 3VSR_A | 2.22e-199 | 76 | 583 | 8 | 488 | Microbacteriumsaccharophilum K-1 beta-fructofuranosidase catalytic domain [Microbacterium saccharophilum],3VSS_A Microbacterium saccharophilum K-1 beta-fructofuranosidase catalytic domain complexed with fructose [Microbacterium saccharophilum] |
| 3WPU_A | 1.11e-198 | 76 | 583 | 8 | 488 | Full-lengthbeta-fructofuranosidase from Microbacterium saccharophilum K-1 [Microbacterium saccharophilum],3WPU_B Full-length beta-fructofuranosidase from Microbacterium saccharophilum K-1 [Microbacterium saccharophilum] |
| 3WPV_A | 2.55e-197 | 76 | 583 | 8 | 488 | Microbacteriumsaccharophilum K-1 beta-fructofuranosidase mutant T47S/F447V/F470Y/P500S [Microbacterium saccharophilum],3WPV_B Microbacterium saccharophilum K-1 beta-fructofuranosidase mutant T47S/F447V/F470Y/P500S [Microbacterium saccharophilum],3WPV_C Microbacterium saccharophilum K-1 beta-fructofuranosidase mutant T47S/F447V/F470Y/P500S [Microbacterium saccharophilum] |
| 3WPY_A | 2.55e-197 | 76 | 583 | 8 | 488 | Microbacteriumsaccharophilum K-1 beta-fructofuranosidase mutant T47S/S200T/F447V/P500S [Microbacterium saccharophilum],3WPY_B Microbacterium saccharophilum K-1 beta-fructofuranosidase mutant T47S/S200T/F447V/P500S [Microbacterium saccharophilum],3WPY_C Microbacterium saccharophilum K-1 beta-fructofuranosidase mutant T47S/S200T/F447V/P500S [Microbacterium saccharophilum] |
| 3WPZ_A | 1.03e-196 | 76 | 583 | 8 | 488 | Microbacteriumsaccharophilum K-1 beta-fructofuranosidase mutant T47S/S200T/F447P/F470Y/P500S [Microbacterium saccharophilum],3WPZ_B Microbacterium saccharophilum K-1 beta-fructofuranosidase mutant T47S/S200T/F447P/F470Y/P500S [Microbacterium saccharophilum],3WPZ_C Microbacterium saccharophilum K-1 beta-fructofuranosidase mutant T47S/S200T/F447P/F470Y/P500S [Microbacterium saccharophilum] |
Swiss-Prot Hits download full data without filtering help
| Hit ID | E-Value | Query Start | Query End | Hit Start | Hit End | Description |
|---|---|---|---|---|---|---|
| Q43998 | 2.44e-187 | 73 | 584 | 65 | 544 | Levansucrase OS=Gluconacetobacter diazotrophicus OX=33996 GN=lsdA PE=1 SV=1 |
| P0DJA3 | 2.99e-88 | 94 | 580 | 11 | 400 | Levansucrase OS=Zymomonas mobilis subsp. mobilis (strain ATCC 31821 / ZM4 / CP4) OX=264203 GN=sacB PE=1 SV=1 |
| F8DT26 | 2.99e-88 | 94 | 580 | 11 | 400 | Levansucrase OS=Zymomonas mobilis subsp. mobilis (strain ATCC 10988 / DSM 424 / LMG 404 / NCIMB 8938 / NRRL B-806 / ZM1) OX=555217 GN=sacB PE=3 SV=1 |
| P0DJA4 | 9.49e-87 | 94 | 582 | 7 | 400 | Extracellular sucrase OS=Zymomonas mobilis subsp. mobilis (strain ATCC 31821 / ZM4 / CP4) OX=264203 GN=sacC PE=3 SV=1 |
| F8DT27 | 9.49e-87 | 94 | 582 | 7 | 400 | Extracellular sucrase OS=Zymomonas mobilis subsp. mobilis (strain ATCC 10988 / DSM 424 / LMG 404 / NCIMB 8938 / NRRL B-806 / ZM1) OX=555217 GN=sacC PE=1 SV=1 |
SignalP and Lipop Annotations help
This protein is predicted as SP
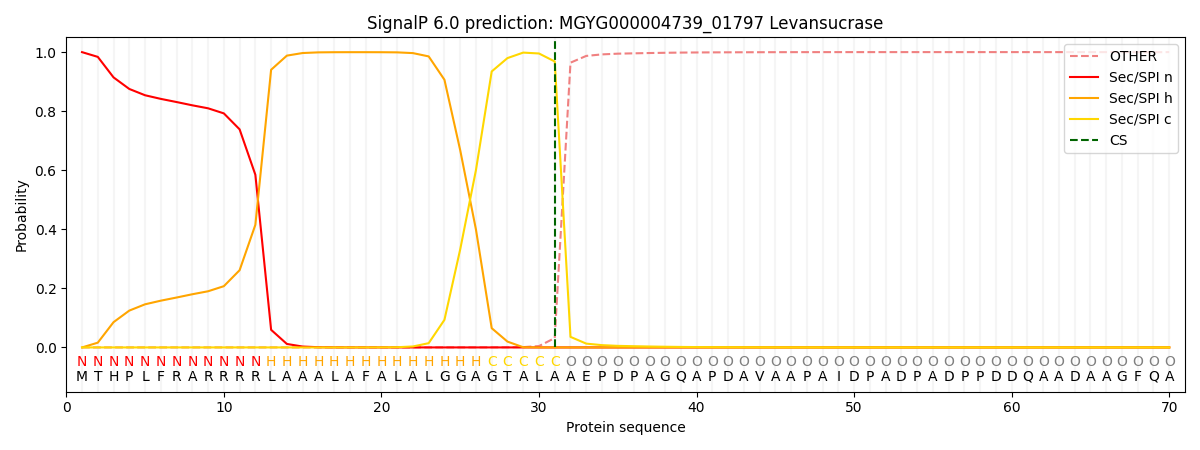
| Other | SP_Sec_SPI | LIPO_Sec_SPII | TAT_Tat_SPI | TATLIP_Sec_SPII | PILIN_Sec_SPIII |
|---|---|---|---|---|---|
| 0.000766 | 0.998187 | 0.000331 | 0.000300 | 0.000219 | 0.000171 |