| Basic Information | |
|---|---|
| Species | Fragaria vesca |
| Cazyme ID | mrna01394.1-v1.0-hybrid |
| Family | GH38 |
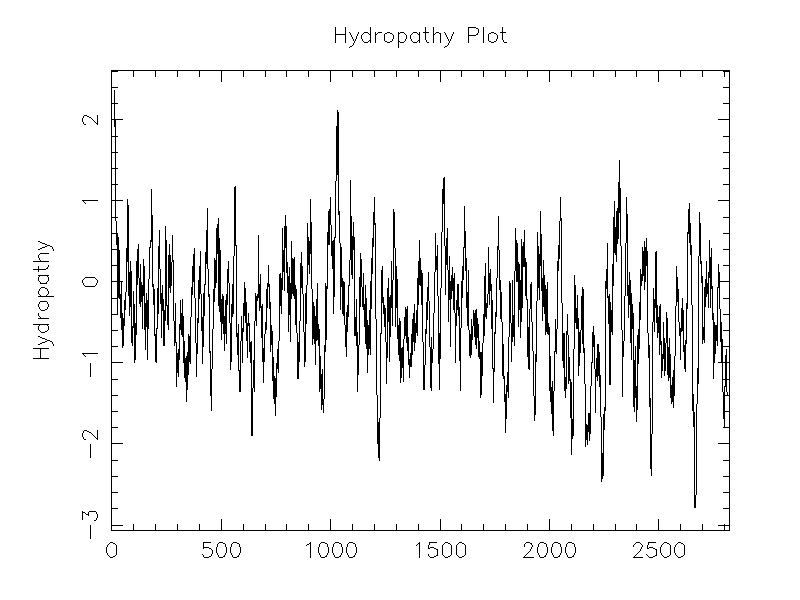
| Protein Properties | Length: 2500 Molecular Weight: 316271 Isoelectric Point: 6.5152 |
| Chromosome | Chromosome/Scaffold: 6 Start: 38026716 End: 38046620 |
| Description | Glycosyl hydrolase family 38 protein |
| View CDS | |
| Signature Domain Download full data set without filtering | |||
|---|---|---|---|
 | |||
| Family | Start | End | Evalue |
| GH38 | 37 | 348 | 0 |
| NVHLVPHSHDDVGWLKTVDQYYVGANNSIRGACVQNVIDSVISSLLEDKNRKFIYVEIAFFQRWWRQQSPALKIKVKELVSSGQLEFINGGMCMHDEATA HYIDLIDQTTLGHQFILKEFGQTPRVGWQIDPFGHSAVQAYLLGAELGFDSLFFARIDYQDRAVRLRDKTLEVIWQGCKSLASSSQIFTGIFPKHYDPPD GFVFEINDVSPPIQDDILLFDYNVQERVNNFVAAALAQANVTRTNHIMWLMGTDFRYQYANSWFRQMDKFIHYVNQDARVNAFYSTPSIYTDAKYAADEQ WPLKTDDFFPYA | |||
| GH38 | 1055 | 1373 | 0 |
| NVHLVPHTHDDVGWLKTVDQYYVGSNNSIQGACVQNVLDSLIPALLADKSRKFVYVEIAFFQRWWREQSDAVQRITKDLVSSGQLEFINGGMCMHDEAAT HYVDMVDQTTLGHRFLKREFNVTPRIGWQIDPFGHSAVQAYLLGAEVGFDSLFFGRIDYQDREKRKNEKSLEFVWRGSKSLGSSAEIFSGAFPENYEPPS GFYFEVNDPSPIDDITLFDYNVQERVNDFIAAAVKQYLTAVIYSQANITRTNHIMWTMGTDFKYQYAHTWFRQMDKLIHYVNKDGRVNALYSTPSIYTDA KYASNESWPIKTDDFFPYA | |||
| Full Sequence |
|---|
| Protein Sequence Length: 2500 Download |
| MASALLLLVV LLAGLLAAQS EYIAYNTTAG IVPEKLNVHL VPHSHDDVGW LKTVDQYYVG 60 ANNSIRGACV QNVIDSVISS LLEDKNRKFI YVEIAFFQRW WRQQSPALKI KVKELVSSGQ 120 LEFINGGMCM HDEATAHYID LIDQTTLGHQ FILKEFGQTP RVGWQIDPFG HSAVQAYLLG 180 AELGFDSLFF ARIDYQDRAV RLRDKTLEVI WQGCKSLASS SQIFTGIFPK HYDPPDGFVF 240 EINDVSPPIQ DDILLFDYNV QERVNNFVAA ALAQANVTRT NHIMWLMGTD FRYQYANSWF 300 RQMDKFIHYV NQDARVNAFY STPSIYTDAK YAADEQWPLK TDDFFPYADH PNAYWTGYFT 360 SRPAFKGYVR ILSGYYLAAR QLEFFKGRSD SGPNTDALAD ALAIVQHHDA VSGTQRQHVA 420 ADYAMRLSIG YLEAEKVVAS SLAYLSDSEP SSGQKRTATK FEQCPLLNIS YCPPSEAALS 480 DGKSLVVVIY NPLGWKREEV IRIPVSYESV TVQDSTQRKI EAQLLPLSNA TLKLRSYYVG 540 AHLGNTPSEP PKYWLAFSVT VPPLGFSSYV ISSAKQTDKD RSSTVSTVYT SEGNTNKTIE 600 VGQGSLKLLY SADEGKLARY INSRYKVTAV AEQSYSYYTG NDGTDKDPQA SGAYVFRPNN 660 TVVIKSEEKV SLTLMRGPVL DEVHQQINQW VSQITRLYKG KEHAEVEFTI GPIPVDDGFG 720 KEITTQFTTA MKTNKTFYTD SNGRDFIKRV RDFRTDWDLQ VNQPIAGNYY PINLGIYVQD 780 SSTELSVLVD RAVGGTSLVD GQVELMLHRR LIHDDIRGVG EVLNETVCIS DKCEGLTVQG 840 KFYLRIDPLG EGSKWRRTAG QEISVPLLLA FAEQEGNDWM NSHVPTFSGI DPSYALPDNV 900 AVITLQELGN GKVLLRLAHL YETGEDKDYS VLANVELKKL FPRKKISKVT ETSLSANQER 960 GEMEKKRLVW KVEGSKESKV VRGGAVDPAK LVVELAPMEI RTFLIDLEYL HIYVRARMAT 1020 KTPGLLFFLA LLLLVADSKF MVYETSQGIV PGKLNVHLVP HTHDDVGWLK TVDQYYVGSN 1080 NSIQGACVQN VLDSLIPALL ADKSRKFVYV EIAFFQRWWR EQSDAVQRIT KDLVSSGQLE 1140 FINGGMCMHD EAATHYVDMV DQTTLGHRFL KREFNVTPRI GWQIDPFGHS AVQAYLLGAE 1200 VGFDSLFFGR IDYQDREKRK NEKSLEFVWR GSKSLGSSAE IFSGAFPENY EPPSGFYFEV 1260 NDPSPIDDIT LFDYNVQERV NDFIAAAVKQ YLTAVIYSQA NITRTNHIMW TMGTDFKYQY 1320 AHTWFRQMDK LIHYVNKDGR VNALYSTPSI YTDAKYASNE SWPIKTDDFF PYADRINAYW 1380 TGYFTSRPAL KHYVRTMSGY YLCNDIFRLT YLYGLLDNKA ARQLEFFKGR SNSGPNTDSL 1440 ADALAIAQHH DAVTGTEKQH VANDYAKRLS IGYMESEELV ATSLAHLVES ASGNPTTKFQ 1500 QAKNLEPLSL CACVLLNISY CPSSEVTLSQ GKKLIVVVYN SLGWKRDDTV RIPVVNEAII 1560 VHDSEGKEVK SQLLPLDDVH LGLRNHYVKA YLGQFPNKTP KYWLVFTVSV PPLGFNTYTI 1620 SGAEGAGAST TRSSVYTIQS KEKSTIEVGQ GNVKLTFSTD EGKMINYVNS RSLVEESVQQ 1680 SYSFYNGYNG SYDKPPLIPQ NAGAYIFRPN GSSFVTPEEK VPLTVMQGPV IDEVHQQINS 1740 WIHQVTRLHK EKEHVEVEFI VGPIPINDGI GKEVVTQLNT TMATNKTFYT DSNGRDFIKR 1800 IRDYRTDWDF KVNQPVAGNY YPINLGIYMQ DDKKEFSVLV DRSLGGSSIV DGQIELMLHR 1860 RLLLDDSRGV AEALNETVCV SDECTGLRVL GKLYFRIDPL GDGAKWRRSF GQEIYSPLLL 1920 AFTEQDGDDW KNSHETTFSG IDSSYSLPDN VALITLQELD DGKVLIRLAH LYEVGEDKDF 1980 SVMANVELKK LFPRKKIGKV TEMNLSANQE RTDMERKRLV WKAEEGSSGT AKGVRGGPVD 2040 PAKLVVELAP MEIRTFLIDL EQNLQRHHYH NFMSILPNQN QGSNSSSSSI PFANPNSQHG 2100 NHHDRHLFQP QSLSRQNIAF GSLDISDSHS PTSPGPSLPA PQDSLGSSQE VTELGLADNN 2160 VMPNHERHSQ SRSGGNGRGA GSYRSRGKRS GSQQSIGTVA SHGSSPQPAG RRSQMMNGNH 2220 LLNFHYDPIS RPQARGPPPP PARRQHKRKA YNKDLFLQAN YKFVVLDAGN YSAESMDPDK 2280 MLRWEDIICV RYSTPTLVQC PICLEYPLCP QITSCGHIFC FPCILQYLLM GKEDHKGDSW 2340 KRCPLCFVMI SPKDLYTLFV ENVKQYSVGD STEFMLLSRQ KDSFTISHKI KQEKDAMPGC 2400 DDESYDPFSK FTFTSDVDLS VRKAISELDA WLVRADAGLV DDLEKLPYVC AAMEHLEQRK 2460 KYWNEHRAID NNKTGSSVVL ATANPTNGNS EASIIGDGTL |
| Functional Domains Download unfiltered results here | ||||||||
|---|---|---|---|---|---|---|---|---|
 | ||||||||
| Cdd ID | Domain | E-Value | Start | End | Length | Domain Description | ||
| PLN02701 | PLN02701 | 2.0e-97 | 1053 | 1872 | 905 | + alpha-mannosidase | ||
| cd00451 | GH38N_AMII_euk | 2.0e-103 | 1054 | 1337 | 284 | + N-terminal catalytic domain of eukaryotic class II alpha-mannosidases; glycoside hydrolase family 38 (GH38). The family corresponds to a group of eukaryotic class II alpha-mannosidases (AlphaMII), which contain Golgi alpha-mannosidases II (GMII), the major broad specificity lysosomal alpha-mannosidases (LAM, MAN2B1), the noval core-specific lysosomal alpha 1,6-mannosidases (Epman, MAN2B2), and similar proteins. GMII catalyzes the hydrolysis of the terminal both alpha-1,3-linked and alpha-1,6-linked mannoses from the high-mannose oligosaccharide GlcNAc(Man)5(GlcNAc)2 to yield GlcNAc(Man)3(GlcNAc)2 (GlcNAc, N-acetylglucosmine), which is the committed step of complex N-glycan synthesis. LAM is a broad specificity exoglycosidase hydrolyzing all known alpha 1,2-, alpha 1,3-, and alpha 1,6-mannosidic linkages from numerous high mannose type oligosaccharides. Different from LAM, Epman can efficiently cleave only the alpha 1,6-linked mannose residue from (Man)3GlcNAc, but not (Man)3(GlcNAc)2 or other larger high mannose oligosaccharides, in the core of N-linked glycans. Members in this family are retaining glycosyl hydrolases of family GH38 that employs a two-step mechanism involving the formation of a covalent glycosyl enzyme complex. Two carboxylic acids positioned within the active site act in concert: one as a catalytic nucleophile and the other as a general acid/base catalyst. | ||
| PLN02701 | PLN02701 | 2.0e-106 | 34 | 1008 | 1090 | + alpha-mannosidase | ||
| cd10810 | GH38N_AMII_LAM_like | 1.0e-168 | 1054 | 1337 | 288 | + N-terminal catalytic domain of lysosomal alpha-mannosidase and similar proteins; glycoside hydrolase family 38 (GH38). The subfamily is represented by lysosomal alpha-mannosidase (LAM, Man2B1, EC 3.2.1.114), which is a broad specificity exoglycosidase hydrolyzing all known alpha 1,2-, alpha 1,3-, and alpha 1,6-mannosidic linkages from numerous high mannose type oligosaccharides. LAM is expressed in all tissues and in many species. In mammals, the absence of LAM can cause the autosomal recessive disease alpha-mannosidosis. LAM has an acidic pH optimum at 4.0-4.5. It is stimulated by zinc ion and is inhibited by cobalt ion and plant alkaloids, such as swainsonine (SW). LAM catalyzes hydrolysis by a double displacement mechanism in which a glycosyl-enzyme intermediate is formed and hydrolyzed via oxacarbenium ion-like transition states. A carboxylic acid in the active site acts as the catalytic nucleophile in the formation of the covalent intermediate while a second carboxylic acid acts as a general acid catalyst. The same residue is thought to assist in the hydrolysis (deglycosylation) step, this time acting as a general base. | ||
| cd10810 | GH38N_AMII_LAM_like | 3.0e-170 | 36 | 312 | 279 | + N-terminal catalytic domain of lysosomal alpha-mannosidase and similar proteins; glycoside hydrolase family 38 (GH38). The subfamily is represented by lysosomal alpha-mannosidase (LAM, Man2B1, EC 3.2.1.114), which is a broad specificity exoglycosidase hydrolyzing all known alpha 1,2-, alpha 1,3-, and alpha 1,6-mannosidic linkages from numerous high mannose type oligosaccharides. LAM is expressed in all tissues and in many species. In mammals, the absence of LAM can cause the autosomal recessive disease alpha-mannosidosis. LAM has an acidic pH optimum at 4.0-4.5. It is stimulated by zinc ion and is inhibited by cobalt ion and plant alkaloids, such as swainsonine (SW). LAM catalyzes hydrolysis by a double displacement mechanism in which a glycosyl-enzyme intermediate is formed and hydrolyzed via oxacarbenium ion-like transition states. A carboxylic acid in the active site acts as the catalytic nucleophile in the formation of the covalent intermediate while a second carboxylic acid acts as a general acid catalyst. The same residue is thought to assist in the hydrolysis (deglycosylation) step, this time acting as a general base. | ||
| Gene Ontology | |
|---|---|
| GO Term | Description |
| GO:0004553 | hydrolase activity, hydrolyzing O-glycosyl compounds |
| GO:0004559 | alpha-mannosidase activity |
| GO:0005975 | carbohydrate metabolic process |
| GO:0006013 | mannose metabolic process |
| GO:0008270 | zinc ion binding |
| Annotations - PDB Download unfiltered results here | |||||||
|---|---|---|---|---|---|---|---|
| Source | Hit ID | E-Value | Query Start | Query End | Hit Start | Hit End | Description |
| PDB | 1o7d_A | 0 | 24 | 318 | 2 | 298 | C Chain C, The Structure Of The Bovine Lysosomal A-Mannosidase Suggests A Novel Mechanism For Low Ph Activation |
| PDB | 1o7d_A | 0 | 1043 | 1343 | 3 | 298 | C Chain C, The Structure Of The Bovine Lysosomal A-Mannosidase Suggests A Novel Mechanism For Low Ph Activation |
| PDB | 1ps3_A | 0 | 12 | 955 | 57 | 1015 | A Chain A, Golgi Alpha-mannosidase Ii In Complex With Kifunensine |
| PDB | 1ps3_A | 0 | 1053 | 1874 | 80 | 882 | A Chain A, Golgi Alpha-mannosidase Ii In Complex With Kifunensine |
| PDB | 3eju_A | 0 | 12 | 955 | 57 | 1015 | A Chain A, Golgi Alpha-mannosidase Ii In Complex With Kifunensine |